
Rodney M. Dale
|
Associate Professor
Ph.D. 2007, University of Chicago
Developmental Biology & Bioinformatics
Phone: 773.508.3606
Fax: 773.508.3646
E-mail: rdale1@luc.edu
|

RESEARCH INTERESTS
My research interests lie in understanding the genetic regulation of critical structural genes and how we can use identified regulatory elements to drive gene expression at desired times and places. My experimental paradigm uses the zebrafish, Danio rerio, to understand the genetic regulation of cartilage and bone formation of the vertebrate skull and notochord. As most vertebrates share a common developmental path using homologous genes and genetic networks to produce a mature animal, the zebrafish has proven to be an excellent tool to study aspects of normal and impaired vertebrate maturation. This is due to the zebrafish being amenable to the powerful and innovative combination of genetic, embryological, molecular, and high-throughput techniques.
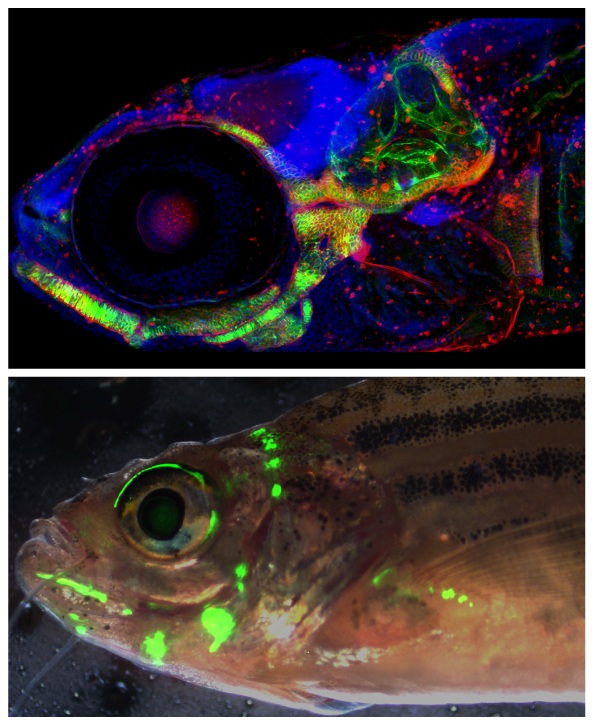
CURRENT PROJECTS:
1. Identification of the tissue specific transcription factor binding sites critical for the col2a1a R2 regulatory element expression.
In Dale and Topczewski 2011, we identified a small novel regulatory sequence (R2) upstream of the col2a1a gene, one of two zebrafish homologues of the mammalian col2a1 gene. The R2 fragment is able to drive gene expression that recapitulates much of the endogenous expression of col2a1a in the developing cartilage, notochord, and ear. The goal of this project will be to genetically dissect the R2 sequence to identify the critical transcription factor binding sites that allow it to drive expression in these three tissue types.
2. Elucidate the mechanism of perichondrial expression of col2a1b during craniofacial development.
Unlike terrestrial vertebrates, the zebrafish, along with other teleost and jawless fish, have two COL2A1 homologues, col2a1a, and col2a1b, suggesting that terrestrial vertebrates lost one of these genes after their evolution from fish. Interestingly, the genomic neighborhood around zebrafish col2a1b has been linked to the human chromosomal region containing COL2A1, and their respective proteins are more closely related than to col2a1a, but based on our previous work the expression pattern of col2a1b is not seen in chondrocytes but in the perichondrium, whereas col2a1a is expressed in both the perichondrium and chondrocytes.
Little is known about the function or the genetic regulation of zebrafish col2a1b, but its expression in the perichondrium around the cartilage elements of the zebrafish cranium makes it an excellent target for promoter analysis for regulatory elements to drive gene expression in the forming perichondrium. While the perichondrium plays a critical part in cartilage development, few have looked at its role specifically. The col2a1b perichondrium regulatory element will allow us to study the morphogenetic changes in the perichondrium during the maturation of the vertebrate animal, which up to this point has not been well characterized.
3. The role of notochord sheath cells in intervertebral disc formation.
Besides congenital defects wear and tear of skeletal elements over a lifetime has become a major healthcare issue, as seen in intervertebral disc (IVD) degeneration and herniation, which are becoming more common in our aging industrialized societies. In mice, it has been postulated that the notochord gives rise to a subpopulation of IVD cells, but how these cells migrate and form this critical structure has yet to be understood. By understanding the process of IVD formation researchers may discover novel therapies for IVD degeneration.
The larval zebrafish notochord is composed of two cell types, vacuolated cells (VC) and notochord sheath cells (NSC). In Dale and Topczewski, 2011, we used our Tg(1.7c2a1a:GFP) reporter fish line to identify the process by which the NSC retracts from the forming embryonic notochord and surrounds the swelling VCs. The vertebrae of the zebrafish spine, called centra, form around the notochord in regular intervals, which are separated by IVD. In our R2 col2a1a derived transgenic fish the NSCs surrounding the notochord where centra are forming loose reporter activity, while those in the region that will give rise to the IVDs maintain reporter activity through adulthood. We are currently setting out to characterize the cellular changes and migration of NSCs play in zebrafish IVD development using our Tg(R2c2a1a:GFP) zebrafish transgenic.
REPRESENTATIVE PUBLICATIONS
Selected peer-reviewed publications
Sisson, BE, Dale, RM, Mui, SR, Topczewska, JM, and, Topczewski, J. A role of glypican4 and wnt5b in chondrocyte stacking underlying craniofacial cartilage morphogenesis. Mechanisms of Development 138:279-290 (2015) https://doi.org/10.1016/j.mod.2015.10.001
Askary, A., Mork, L., Paul, S., He, X., Izuhara, A. K., Gopalakrishnan, S., Ichida, J. K., McMahon, A. P., Dabizljevic, S., Dale, R., Mariani, F. V., Crump, J. G. Iroquois Proteins Promote Skeletal Joint Formation by Maintaining Chondrocytes in an Immature State. Developmental Cell. 35:358-365 (2015) https://doi.org/10.1016/j.devcel.2015.10.004
Gray, R. S., Wilm, T. p., Smith, J., Bagnat, M., Dale, R., Topczewski, J., Johnson, S. L., Solnica-Krezel, L. Loss of col8a1a function during zebrafish embryogenesis results in congenital vertebral malformations. Developmental Biology. 386:72-85 (2014) https://doi.org/10.1016/j.ydbio.2013.11.028
Antoine, T. E., Jones, K. S., Dale, R., Shukla, D., Tiwari, V. Zebrafish: Modeling for Herpes Simplex Virus Infections. Zebrafish 11:17-25. (2014) https://doi.org/10.1089/zeb.2013.0920
Topczewski, J, Dale, RM, and Sisson, BE. PCP signaling in Craniofacial Development, Organogenesis, 7:255-59. (2011) http://dx.doi.org/10.4161/org.7.4.18797
Dale, RM, and Topczewski, J, Identification of an evolutionarily conserved regulatory element of the zebrafish col2a1a gene, Developmental Biology, Vol. 357, (2):518-531. (2011) http://doi.org/10.1016/j.ydbio.2011.06.020
Demonbreun, AR, Lapidos, KA, Heretis, K, Levin, S, Pytel, P, Dale, RM, Svensson, EC, McNally, EM. Myoferlin regulation by NFAT in muscle injury, regeneration, and repair. Journal of Cell Science, 123:2413-22. (2010) https://doi.org/10.1242/jcs.065375
Dale, RM, Sisson, BE, and Topczewski, J, The emerging role of Wnt/PCP signaling in organ formation, Zebrafish, 6:9-14. (2009) https://doi.org/10.1089/zeb.2008.0563
Dale, RM, Remo, BF, and Svensson, EC. An alternative transcript of the FOG-2 gene encodes a FOG-2 isoform lacking the FOG repression motif, Biochemical and Biophysical Research Communications, Vol. 357 (3): 683-687. (2007) https://doi.org/10.1016/j.bbrc.2007.04.008
Book Chapters
Svensson, EC, Wilk, J, Dale, RM, and Modrell, M. “The role of the transcriptional co-repressor FOG-2 in cardiac development” in, Cardiovascular Development and Congenital Malformations Artman, M., Benson, D.W., Srivastava, D., and Nakazawa, M. (eds), p125-127. (2008)